

Immunoglobulins In Poultry
BiRan DaiManman Caoweisheng LiaoMing (College of Agriculture,South China Agricultural University, Guangdong Guangzhou 510642)
Abstract:Immunoglobulin is a kind of globulin which exists in human, animal serum, tissue fluid and other exocrine fluids, which is the essence of antibody. At present, there are three kinds of immunoglobulins in avian including IgY, IgM and IgA, which have different functions and play an important role in the immune system. In this paper, the structure and function, biological effects and production rules of avian immunoglobulin are briefly introduced.
Key words:immunoglobulin,poultry,IgA,IgY, IgM
1、前言
免疫球蛋白(immunoglobulins)是存在于人和动物血清、组织液及其他外分泌液中的一类具有抗体性质的球蛋白。在禽类中已发现 IgY、 IgM 和 IgA,尚未观察到 IgD 和 IgE。禽类抗体虽然与哺乳动物抗体相似,但具有显著不同的生化功能和特性,这些特性为研究人员在免疫诊断和免疫治疗的开发与应用方面提供了更具体 的参考。
2、禽免疫球蛋白的基本结构
所有免疫球蛋白单体的结构都是由两条重 链和两条轻链组成的“Y”型结构[1]。两条重链之间和重链与轻链羧基端(C 端)之间以二硫键相连。重链氨基端(N 端)开始最初的110 个氨基酸的排列顺序以及结构是随抗体分子的特异性而变化,这一区域称为重链的可变区(VH),其余的氨基酸比较稳定,称为恒定区(CH)。轻链 从氨基酸开始最初的 109 个氨基酸(约占轻链 的 1/2)称为轻链的可变区(VL),其余的称为恒定区(CL)。免疫球蛋白的多肽链分子可折叠成几个环状球形结构,这些结构被称为免疫球蛋白的功能区,每个功能区大概由 110 个氨基酸 组成。轻链有两个功能区,即 VL 和 CL,分别位于可变区和恒定区,禽类的重链普遍有 5 个功能区,其中一个功能区(VH)在可变区,其余的 (CH1、CH2、CH3、CH4)在恒定区[2-4]。此外,哺乳动物 CH1 与 CH2 之间有一段氨基酸序列称为铰链区,与抗体分子的构型变化有关,但在家禽免疫球蛋白中并没有类似的铰链区。从进化的观点来看,哺乳动物免疫球蛋白 CH 的铰链区起源于禽免疫球蛋白 CH 中 CH2 结构域的缩合[5-7]。Porter(1959)用木瓜蛋白酶水解 IgG,得到 三个片段,其中两个相同片段称为抗原结合片段(Fab),由一条完整的轻链和N 端 1/2 重链组成。另一个片段可形成蛋白结晶,称为 Fc 片段, 该片段虽无结合抗原的活性,但具有各类免疫球蛋白的抗原决定簇,并与抗体分子的其他生物学活性有密切关系。依据重链的理化特性及抗原差异,可将禽鸟体内的免疫球蛋白分为 IgM、IgA、IgY(IgG),之前普遍认为在禽鸟体内不存在 IgD[8],然而最近基于鸟类基因组测序和免疫相关组织的转录组测序,证明了鸵鸟(Struthio camelus)具有编码膜结合的 IgD 重链的功能性 δ 基因,这赋予家禽免疫球蛋白研究新的启发[9]。同一种免疫球蛋白,又可根据重链恒定区细微结构分为亚型,如鸡的 IgY 可分为 IgY1、IgY2、IgY3 亚型,鸭的 IgY 可分为 IgY1 (7.8S)和 IgY2(5.7S)。根据轻链恒定区的不同可将轻链分为 λ、k 两个亚型,禽类的轻链表现为 λ 亚型[10]。
3、各类免疫球蛋白特征及功能
利用免疫细胞化学和遗传学技术,已在家禽中鉴定出三类免疫球蛋白:IgY、IgM 和 IgA, 其分子结构及其体内特征各不相同,如表 1。
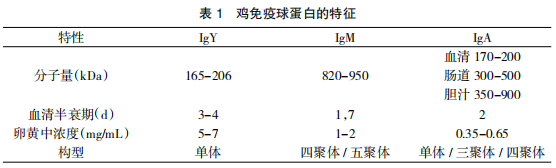
3.1 IgY
禽类 IgY 为单体免疫球蛋白,从系统发育学上来看与哺乳动物免疫球蛋白相距甚远,因此具有非常不同的特性。1969 年,Gerrie Leslie 和 Bill Clem 提出,鸡的主要血清免疫球蛋白应称为 IgY,而不是 IgG。也有研究表明哺乳动物 IgG 和 IgE 可能是由禽类 IgY 分化而来 [11]。鸡 IgY 分子量大,约为 16.5~20.6 万道尔顿,含糖量高达 6%(约为人 IgG 的两倍),沉降系数为 7.8S。然而在鸭[12]等雁形目中存在两种一大一小 的 IgY“异构体”,其中较大的 IgY“异构体”在结构上类似于鸡的 7.8S IgY,较小的“5.7S IgY”被 称为 IgY(AFc),它的 H 链只有三个结构域(V、 HC1 和 HC2),在结构和抗原上类似于 IgY 的 F (ab’)2 片段。在 IgY 中,有一段特殊的区域,含有脯氨酸和甘氨酸残基,这些区域有可能赋予 IgY 一定的“灵活性”,类似于哺乳动物免疫球蛋白的铰链区。IgY 的“灵活性”程度可以解释一些独特的生化特性,例如 IgY 通常能牢固地结合抗原,只有在高浓度盐水下才能引起沉淀或凝集反应。还原剂和蛋白水解酶可以异常降解家禽 IgY,这对家禽 IgY 的结构研究造成一定的困难,但是家禽 IgY 具有较强的耐热、耐酸和抗酶解能力[13]。
在受到抗原刺激时,禽类可以将血清中的 IgY 选择性地转移到卵黄中,因此又将 IgY 命名为卵黄免疫球蛋白。现在的研究越来越强调哺乳动物 IgG 和家禽 IgY 之间的比较,因为他们是基于抗体产生和开发应用中最相关的抗体同种型,比如 IgY 不能结合许多哺乳动物的免疫调节蛋白和补体[14],这可以规避各种免疫干扰,使鸡单克隆抗体越来越多的用于哺乳动物和人的免疫诊断和免疫治疗[15-18]。
3.2 IgM
禽 IgM 在结构和功能上与哺乳动物 IgM 相似,主要分布于血管中,约1~2mg/mL,分子量约为 90 万道尔顿,沉降系数为 17S,含糖量 7%。 然而,H 链和 L 链分子量分别为 7 万道尔顿和 2 万道尔顿,表明鸡 IgM 更有可能是四聚体结构而不是五聚体,或者可能是两者的混合物[19]。
IgM 是初次免疫应答最早产生的抗体,所 以在早期诊断中具有重要意义。IgM 具有多个抗原结合位点,有抗细菌、抗病毒和中和毒素等免疫活性,是一种高效的抗体。由于 IgM 分子在免疫应答中的进化保守性及其中和作用短暂的特点,禽类 IgM 生物学的研究受到一定限制。
3.3 IgA
1972 年,在鸡的血清和分泌物中发现了第三类免疫球蛋白 IgA[20]。IgA 以单体和二聚体两种分子形式存在,单体存在于血清中,含量较低。二聚体为分泌型的 IgA(SIgA),由呼吸道、消化道、泌尿生殖道等部位的黏膜固有层中的浆细胞所产生。两个单体由一条 J 链连接在一 起形成二聚体,二聚体与分泌性上皮细胞产生的分泌组分(SC)连接以形成 SIgA,SIgA 最终通过分泌性上皮细胞分泌到腺体腔中。SIgA 在胆汁和肠道中的含量最高,分别为 3.5mg/mL 和 1.5mg/ml。鸡 IgA 被确定为胆汁中唯一存在的免疫球蛋白类[21]。
SIgA 作为参与黏膜免疫的重要因素,在防御各种致病性病原(如细菌、病毒和寄生虫)感染中起着关键作用[22]。作为一种相对独立的免疫系统,黏膜免疫系统(MIS)主要由肠黏膜免疫[23]、支气管黏膜免疫、泌尿生殖道黏膜免疫和结膜局部免疫组成。在传染病的预防接种中,经点眼、滴鼻、饮水及气雾途径免疫,均可产生分泌型 IgA,而建立相应的黏膜免疫。现在黏膜免疫也应用于家禽寄生虫的预防[24]。
3.4 缺失的 IgD 和 IgE
IgD 作为原始的免疫球蛋白同种型,已在大多数下颚脊椎动物中被鉴定出来[25]。虽然有鸡的 IgD 同源物[26]的报道,也有发现马血清刺激成年家禽后可能产生IgE[27],但普遍认为禽类没有 IgD 和 IgE[8]。
4、生物效应
抗体分子的生物学活性与 Fc 有关。 Fc 片段在 IgY 从卵黄转运到血液中起关键作用[28,29]。禽类的抗体传递分为两个步骤,第一步是将抗体转移到卵黄,而禽类卵黄薄膜上的 Fc 受体在第二步发挥作用,将卵黄囊内的 IgY 转运至禽胚血液中[30]。且 IgY 与 Fc 受体的结合受酸碱性的影响[31],即 pH=6 时相互结合, pH=7 时解离。当重组 IgY-Fc 时,其转运水平受到影响[32]。
Fc 片段可以决定免球蛋白分子的亲细胞性。一些免疫细胞如巨噬细胞、淋巴细胞、嗜碱性粒细胞、肥大细胞等表面都具有免疫球蛋白 Fc 片段的受体,因此免疫球蛋白可通过其 Fc 片段与这些带有 Fc 受体的细胞结合。免疫球蛋白与这些细胞受体的结合部位因其种类不同而有差异。
Fc 片段还与免疫球蛋白通过黏膜进入外分泌液有关,如分泌型 IgA 可由局部黏膜固有层中的浆细胞产生,然后通过黏膜进入呼吸道和消化道分泌物中,此过程与 Fc 片段有关[33]。Fc 片段是免疫球蛋白的重链稳定区,因此它决 定了各免疫球蛋白的抗原决定性。
5、抗体产生特点
禽类免疫球蛋白均由 B 淋巴细胞产生。骨髓淋巴干细胞经血液流至法氏囊后被诱导分化成 B 淋巴细胞,这些细胞膜上分别带有 SIgY、 SIgM、SIgA 的淋巴细胞离开法氏囊后在盲肠扁桃体、脾脏和其他淋巴组织中定居,受到抗原刺激后即分化为浆细胞并产生抗体,参与体液免疫[34]。有证据表明,在鸡胚法氏囊中,IgM 阳性细胞最早在孵化第 10 天可检测到,至孵化第 13 天已能检测到 IgY 阳性细胞,至孵化第 16 天时能检测到 IgA 阳性细胞[35]。雏鸡出壳后由于受到母源抗体的影响,在 3~5 天龄时一直保持较高的 IgY 水平,但各免疫器官 B 淋巴细胞始终以 IgM 阳性细胞含量多[35]。禽类受到抗原刺激后,最先产生特异性的IgM,然后通过类似的转换生成 IgY 和 IgA。
6、总结
免疫球蛋白是机体免疫系统对抗外界抗原的重要效应分子,然而目前对禽类免疫球蛋白的研究仅局限于鸡形目和雁形目的少数几个物种,发现他们只表达 IgM,IgA,IgY 三种免疫球蛋白重链基因,并且通过基因转换机制产生免疫球蛋白可变区多样性。禽类相对于哺乳动物来说具有独特的免疫系统,理解他们的差异对于家禽和哺乳动物都是有益的。随着分子生物学和高通量测序技术的发展以及各类家禽基因组的不断解析,对于家禽免疫球蛋白的研究更趋向于分子层面,也为免疫球蛋白结构与功能特点及其应用的进一步探索提供了空间。家禽免疫球蛋白的研究不仅可为家禽疾病防控提供新的解决方案,也利于防控人兽(畜)共患病从而保证人类健康,并有助于解决全球食品安全问题。
参考文献
1. Roux K H. Immunoglobulin structure and function as revealed by electron microscopy. Int Arch Allergy Immunol, 1999. 120(2): 85-99.
2. Taylor A I, FabianeS M,Sutton B J, et al.The crystal structure of an avian IgY -Fc fragment re- veals conservation with both mammalian IgG and IgE. Biochemistry, 2009. 48(3): 558-562.
3. Dahan A, ReynaudCA,.WeillJ C. Nucleotide sequence of the constant region of a chicken mu heavy chain immunoglobulin mRNA. Nucleic Acids Res, 1983. 11(16): 5381-5389.
4. Backhed F, Roswall J, Peng Y Q, et al. Dy- namics and Stabilization of the Human Gut Micro- biome during the First Y ear of Life. Cell Host Microbe, 2015. 17(6): 852.
5. Mansikka A. Chicken IgA H chains. Implica-tions concerning the evolution of H chain genes. J Immunol, 1992. 149(3): 855-861.
6. Parvari R, Avivi A,Lentner F, et al. Chicken immunoglobulin gamma-heavy chains: limited VH gene repertoire, combinatorial diversification by D gene segments and evolution of the heavy chain lo- cus. Embo j, 1988. 7(3): 739-744.
7. Choi J W, Kim J K, Seo H W, et al. Molecular cloning and comparative analysis of immunoglobu- lin heavy chain genes from Phasianus colchicus, Meleagris gallopavo, and Coturnix japonica. Vet Immunol, 2010. 136(3-4): 248-256.
8. Magor K E, Navarro D M,Barber M R W, et al. Defense genes missing from the flight division. Dev Comp Immunol, 2013. 41(3): 377-388.
9. Han B Y, Yuan H,Wang T,et al. Multiple IgH Isotypes Including IgD, Subclasses of IgM, and IgY A re Expressed in the Common A ncestors of Mod- ern Birds.J Immunol, 2016. 196(12): 5138-5147.
10. Reynaud C A, DahanA, WeillJ C. Complete sequence of a chicken lambda light chain im-munoglobulin derived from the nucleotide se-quence of its mRNA. Proc Natl Acad Sci U S A, 1983. 80(13): 4099-4103.
11. Warr G W, Magor K E, HigginsD A .IgY: clues to the origins of modern antibodies. Immunol Today, 1995. 16(8): 392-398.
12. Hatta H, Tsuda K, Akachi S, et al. Productivity and some properties of egg yolk antibody (IgY ) a- gainst human rotavirus compared with rabbit IgG. Biosci Biotechnol Biochem, 1993. 57(3): 450-454.
13. Higgins D A, Warr G W.Duck immunoglobu- lins: Structure, functions and molecular genetics. Avian Pathol, 1993, 22(2): 211-236.
14. Lee W,Atif A S, Tan S C, et al. Insights into the chicken IgY with emphasis on the generation and applications of chicken recombinant mono- clonal antibodies. J Immunol Methods, 2017. 447: 71-85.
15. Carlander D,Stalberg J,Larsson A.Chicken an- tibodies: a clinical chemistry perspective. Ups J Med Sci, 1999. 104(3): 179-189.
16. Lanzarini N M., BentesG A.,Volotao E D.Use of chicken immunoglobulin Y in general virology. J Immunoass Immunoch, 2018. 39(3): 235-248.
17. Thu H M.,MyatT W, Win M M, et al. Chicken egg yolk antibodies (IgY ) for prophylaxis and treat- ment of rotavirus diarrhea in human and animal neonates: a concise review. Korean J Food Sci Anim Resour, 2017. 37(1): 1-9.
18. da SilvaW D,Tambourgi D V,IgY: a promising antibody for use in immunodiagnostic and in im- munotherapy. Vet Immunol Immunopathol, 2010. 135(3-4): 173-180.
19 Leslie G A, Clem L W,Phylogeny Of Im- munoglobulin Structure A nd Function .III. Im-munoglobulins Of The Chicken. J Exp Med, 1969, 130(6): 1337.
20. Lebacqve A M, VaermanJ P., Heremans,J.F., A possible homologue of mammalian IgA in chick- en serum and secretions. Immunology, 1972. 22(1): 165-175.
21. Sanders B G., CaseW L,Chicken secretory immunoglobulin: chemical and immunological characterization of chicken IgA. Comp Biochem Physiol B-BiochemMol Biol, 1977. 56 (3): 273-278.
22. Macpherson A J, MccoyK D., Johansen F E., et al., The immune geography of IgA induction and function. Mucosal Immunol, 2008. 1(1): 11-22.
23. Stockinger S, HornefM W, Chassin,C.,Estab- lishment of intestinal homeostasis during the neonatal period. Cell Mol Life Sci, 2011. 68 (22): 3699-3712.
24. Tian E J,Zhou B H., WangX Y, et al., Effect of diclazuril on intestinal morphology and SIgA ex- pression in chicken infected with Eimeria tenella. Parasitol Res, 2014. 113(11): 4057-4064.
25. Ohta Y, Flajnik M,IgD, like IgM, is a primor- dial immunoglobulin class perpetuated in most jawed vertebrates. Proc Natl Acad Sci U S A, 2006. 103(28): 10723-10728.
26. Chen CL H, Lehmeyer J E,Cooper M D,Evi- dence for an IgD homologue on chicken lympho- cytes. J Immunol, 1982.129(6): 2580-2585.
27 Burns R B, Maxwell M H,Probable occurrence of IgE in the adult domestic fowl (Gallus domesti- cus) after horse serum stimulation. Vet Res Commun, 1981.5(1):67-72.
28. Tian Z H, Zhang X. Progress on research of chicken IgY antibody-FcRY receptor combination and transfer. J Recept Signal Transduct, 2012.32 (5):231-237.
29. Higgins D A, Warr G W.Duck immunoglobu- lins: structure, functions and molecular genetics. Avian Pathol, 1993.22(2): 211-236.
30. West A P, Herr A B, Bjorkman P J. The chick- en yolk sac IgY receptor, a functional equivalent of the mammalian MHC -related fc receptor, is a phospholipase A (2) receptor homolog. Immunity, 2004.20(5): 601-610.
31. Linden C D, Roth T F. IgG receptors on fetal chick yolk sac. J Cell Sci, 1978.33(OCT):317-328.
32. Bae H,Honda H,Murota R, et al. Production of recombinant chicken IgY -Fc and evaluation of its transport ability into avian egg yolks. J PoultSci, 2010. 47(3): 256-261.
33. Monteiro R C,Jg V D W. IgA Fc receptors. Annu Rev Immunol, 2003.21:177-204.
34. Ciriaco E,PineraP P,Diaz-Esnal B, et al. A ge – related changes in the avian primary lymphoid or- gans (thymus and bursa of fabricius). Microsc Res Tech, 2004.62(6):482-487.
35. 杨丽华. 鸡法氏囊中 B 淋巴细胞的发育及其免疫功能发生的研究 [D]. 吉林农业大学, 2007.
作者:毕冉 1 代曼曼 1 曹伟胜 1 廖明 1 杨皓 2 (1.华南农业大学兽医学院,广州 510642; 2.默沙东动物保健品(上海)有限公司,上海 200233)





